We've been taking a tour of Mammalia for the past two annual "Fossil [Group] of the National Park Service", and this year we'll make it three in a row with man's best friends, the Canidae. The dog record starts up in the Eocene with the hesperocyonines, who held court through the Oligocene but then petered out in the Miocene. Turning up in the Oligocene are the borophagines, or "bone-crushing dogs," and the canines, which include wolves, foxes, domestic dogs, and close relatives. Borophagines, as the nickname suggests, had robust jaws and teeth, which doesn't mean they should be typecast as slavering hypercarnivorous brutes (canids in general have been pretty flexible about diet over their history). They drop out of the record at about the end of the Pliocene, leaving the field clear for the canines, which had been a fairly minor component of the canid radiation until about the Late Miocene. If you're interested in paleontological nitty-gritty on these two groups, check out Wang et al. (1999) on borophagines and Tedford et al. (2009) on canines. Canids, incidentally, are a North American invention, and unlike some other groups that originated in North America (camels, horses, rhinos), they have never gone extinct on the continent.
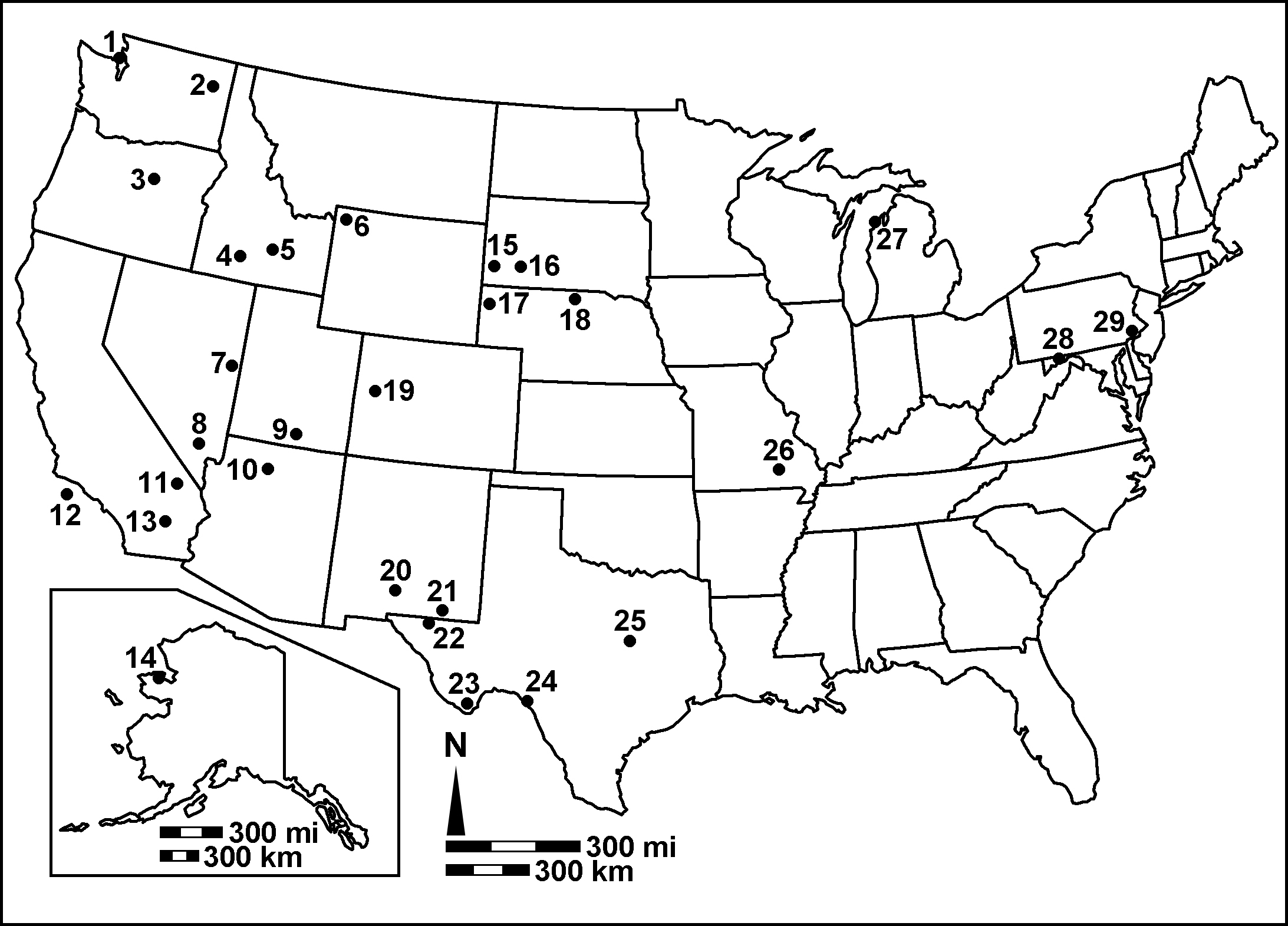
I have records of 29 NPS units with canid fossils, essentially through the mid-Holocene (omitting the gray areas of archeological overlap). The distribution, as seen in the map below, is pretty similar to the distribution of parks with felids and lagomorphs from the past two years. This is entirely unsurprising, especially for the pre-Quaternary records, because those are the parks where we have significant mammal fossils in general. They're a bit less abundant than I would have suspected, but then again canids aren't typically the main show at fossil sites.
|
|
Map time! 1. Ebey's Landing National Historical Reserve; 2. Lake
Roosevelt National Recreation Area; 3. John Day Fossil Beds National
Monument; 4. Hagerman Fossil Beds NM; 5. Craters of the Moon NM and
Preserve; 6. Yellowstone National Park; 7. Great Basin NP; 8. Tule
Springs Fossil Beds NM; 9. Glen Canyon NRA; 10.
Grand Canyon NP; 11. Death Valley NP; 12. Channel Islands NP; 13. Mojave
National Preserve; 14. Bering Land Bridge NPres; 15. Wind Cave NP; 16.
Badlands NP; 17. Agate Fossil Beds NM; 18. Niobrara National Scenic
River; 19. Colorado NM; 20. White Sands NP; 21. Carlsbad Caverns NP; 22.
Guadalupe Mountains NP; 23. Big Bend NP; 24. Amistad NRA; 25. Waco
Mammoth NM; 26. Ozark National Scenic Riverways; 27. Sleeping Bear Dunes
National Lakeshore; 28. Potomac Heritage National Scenic Trail; 29.
Valley Forge National Historical Park. |
Of that 29, most are Quaternary records. Eight parks have pre-Quaternary records: Agate Fossil Beds NM, Badlands NP, Big Bend NP, Hagerman Fossil Beds NM, John Day Fossil Beds NM, Mojave NPres, Niobrara NSR, and Wind Cave NP. Six of those are completely unsurprising, but you'd have to be familiar with the geology of Mojave and Wind Cave to guess those two. Caves at Carlsbad NP, Potomac Heritage NST, and Valley Forge NHP chip in three more ancient Pleistocene records, but after that you're looking at the past 50,000 years. Below is a figure here showing distribution through time. Instead of the usual "Late Eocene, Early Oligocene", etc., I did them in North American Land Mammal Ages; I'll give brief approximate translations in the caption. (If you're not familiar with NALMAs, this is great stuff to know if you want to hang out with mammal paleontologists.)

|
| Click to embiggen. It's best not to get hung up on the exact dates, as they tend to vary from source to source (the relative dates of biostratigraphy not always playing nice with chronometric dating from things like ash). Chadronian ≈ 37–33.9 Ma (million years ago), Late Eocene; Orellan ≈ 33.9–31.8 Ma, Early Oligocene; Whitneyan ≈ 31.8–29.5 Ma, Early Oligocene; Arikareean ≈ 29.5–18.5 Ma, Early Oligocene–Early Miocene; Hemingfordian ≈ 18.5–16.3 Ma, Early Miocene; Barstovian ≈ 16.3–12.5 Ma, Early–Middle Miocene; Clarendonian ≈ 12.5–9.4 Ma, Middle–Late Miocene; Hemphillian ≈ 9.4–4.7 Ma, Late Miocene–Early Pliocene; Blancan ≈ 4.7–1.4 Ma, Early Pliocene–Early Pleistocene; Irvingtonian ≈ 1.4–0.21 Ma, Early–Middle Pleistocene. |
It's not a bad distribution; between eleven parks, most of the featured NALMAs have at least two representatives (the Arikareean gets a helping hand by being at least twice as long as the others). The broadest records belong to Badlands, John Day, and Niobrara, which together give a view of canids from the beginning of the Oligocene to the end of the Miocene. Significant recent information can be found on the paleontology of Badlands and John Day in Benton et al. (2015) and Fremd (2010), respectively, although do note that both publications have a wider scope than just the areas within park boundaries. Niobrara takes a bit more work, but if you've got Skinner and Johnson (1984), you've got a lot of the story, and if you've got Voorhies (1990; easier said than done!), you've got about everything. Badlands has about ten genera in the White River Group (Benton et al. 2015; I did not include the Chadronian record of Hesperocyon due to it being well outside of the park, but you get the idea). In the John Day Basin, the Turtle Cove Member of the John Day Formation has yielded 13 genera, although they were not all contemporaries; higher units are not as extravagant but still have two to five genera (Fremd 2015). At Niobrara, most of the Valentine Formation sites have one or more of the borophagines Aelurodon and Paratomarctus and the early canine Leptocyon, and a couple other borophagines are present at Norden Bridge Quarry (Voorhies 1990). There's only one strong canid site in the overlying Ash Hollow Formation, Connection Kat Quarry, but it is similar to Norden Bridge with Leptocyon (Tedford et al. 2009) and five borophagines (Wang et al. 1999).
By the time we get to the Quaternary, the number of genera is much reduced, with just Aenocyon (resurrected for the dire wolf in 2021 on molecular evidence), Canis, Urocyon, and Vulpes. Generally speaking, most records at the species level are of gray wolf (Canis lupus), coyote (Canis latrans), gray fox (Urocyon cinereoargenteus), and red fox (Vulpes vulpes), often more than one of the four at a site. The extinct Armbruster's wolf (Aenocyon or Canis armbrusteri) shows up at the oldest Pleistocene sites (Carlsbad, Potomac Heritage, and Valley Forge), and the ever-popular dire wolf (Aenocyon dirus) is reported from four park units (Carlsbad, Ozark, Tule Springs, and Wind Cave).

|
|
Urocyon littoralis, watching us drive by on Santa Rosa Island
(Channel Islands NP). This species is historically thought of as a
Pleistocene arrival, but reliable records only go back to the
mid-Holocene (Rick et al. 2009). |
For these Fourth of July posts, I usually list the fossil species named from specimens found in or potentially found in NPS areas. In this case, I have four definite and 20 potential names; I thought it might be tedious to list the 20 potentials, so I'm just going to list the four definites:
- Canis armbrusteri Gidley (1913) from Cumberland Bone Cave, Potomac Heritage NST
- Canis priscolatrans Cope (1899) from Port Kennedy Bone Cave, Valley Forge NHP (basically the same as Canis edwardii but based on scrappy material)
- Nothocyon annectens Peterson (1907) from Agate Fossil Beds NM (now Phlaocyon annectens)
- Oxetocyon cuspidatus Green (1954) from Badlands NP
Armbruster's wolf is probably the most familiar, due to its not-too-distant Pleistocene age and association with the dire wolf.
References
Benton, R. C., D. O. Terry, Jr., E. Evanoff, and H. G. McDonald. 2015. The White River Badlands: geology and paleontology. Indiana University Press, Bloomington and Indianapolis, Indiana.
Cope, E. D. 1899. Vertebrate remains from Port Kennedy bone deposit. Journal of the Academy of Natural Sciences of Philadelphia, 2nd series, 11(3):193–286.
Fremd, T. J. 2010. SVP Field Symposium 2010: John Day Basin Field Conference. Society of Vertebrate Paleontology.
Gidley, J. W. 1913b. Preliminary report on a recently discovered Pleistocene cave deposit near Cumberland, Maryland. Proceedings of the United States National Museum 46:93–102.
Green, M. 1954. A cynarctine from the upper Oligocene of South Dakota. Transactions of the Kansas Academy of Science 57:218–220.
Peterson, O. A. 1907 [sometimes given as 1906]. The Miocene beds of western Nebraska and eastern Wyoming and their vertebrate faunae. Annals of the Carnegie Museum 4:21–72.
Rick, T. C., J. M. Erlandson, R. L. Vellanoweth, T. J. Braje, P. W. Collins, D. A. Guthrie, and T. W. Stafford, Jr. 2009. Origins and antiquity of the island fox (Urocyon littoralis) on California's Channel Islands. Quaternary Research 71:93–98.
Skinner, M. F., and F. W. Johnson. 1984. Tertiary stratigraphy and the Frick Collection of fossil vertebrates from north-central Nebraska. American Museum of Natural History Bulletin 178:215–368.
Tedford, R. H., X. Wang, and B. E. Taylor. 2009. Phylogenetic systematics of the North American fossil Caninae (Carnivora: Canidae). Bulletin of the American Museum of Natural History 325.
Voorhies, M. R. 1990. Vertebrate paleontology of the proposed Norden Reservoir Area, Brown, Cherry, and Keya Paha counties, Nebraska. Division of Archeological Research, University of Nebraska, Lincoln, Nebraska. Technical Report 82-09.
Wang, X., R. Tedford, and B. E. Taylor. 1999. Phylogenetic systematics of the Borophaginae (Carnivora, Canidae). Bulletin of the American Museum of Natural History 243.
No comments:
Post a Comment