The joys of Lameta titanosaurs, or: Lameta Lamentations
A brief introduction to the titanosaurs of the Lameta Formation is in order, because they're all tied together in one big figurative "titanosaur king". The Lameta titanosaurs have historically been messy, they're messy today, and they'll be messy in the future. It's just the nature of the game when you're dealing with a situation where 1) fossils have been reported since the late 1820s (Matley 1921; Wilson and Upchurch 2003), 2) quarry maps frequently weren't made or no longer exist, 3) associated specimens are uncommon anyway, 4) museum specimens have gone missing, and 5) Friedrich von Huene got to work with a bonebed. At the moment, most people (with Malkani as an exception) seem satisfied with a gracile species (Isisaurus colberti) and a robust species (Jainosaurus septentrionalis), with room for possible additions such as whatever produced the Nand axis (Wilson and Mohabey 2006), but check back again in a few years, or if someone describes another associated specimen.I kinda buried part of the lede with my brief mention of Malkani's recent synonymizations; they also extended to the established Lameta titanosaurs (Malkani 2019a, 2019b). He proposed that Titanosaurus indicus was the same as Pakisaurus balochistani, as well as possibly the holotype braincase of Jainosaurus septentrionalis (regarding the postcrania associated with the type of Jainosaurus as chimeric); and that Titanosaurus blanfordi and the Indian material of Titanosaurus madagascariensis pertain to Saraikimasoom vitakri. A variety of questions come to mind, not least of which being "if the types of T. indicus and T. blanfordi are diagnostic enough to be considered for synonymization, shouldn't they then be the taxa of record based on priority?" It's not as critical with Malkani's own species, if you agree with some commentators that none of them are official by the rules of the ICZN and thus none would have priority over any of others, but the species of Titanosaurus have unquestionable priority over almost every other titanosaur species. By priority, his foursome of Isisaurus colberti, Pakisaurus balochistani, Saraikimasoom vitakri, and Gspsaurus pakistani would become Isisaurus colberti, Titanosaurus indicus, Saraikimasoom blanfordi, and Gspsaurus pakistani.
Isisaurus colberti
Isisaurus colberti is the easy one to deal with. It's got the only unquestionably associated material and it was first described in the 1990s, so it doesn't have an interminable paper trail behind it. Despite the good material, it has only been the primary subject of a couple of papers: Jain and Bandyopadhyay (1997), which named it (as Titanosaurus colberti), and Wilson and Upchurch (2003), which moved it to its present genus. You might also count Sharma et al. (2005) if you're the kind of person who is confident about knowing the producers of coprolites to the genus level, but I'm not. Otherwise, it's a lot of asides and mentions. Like the old saw about the weather, people talk a lot about Isisaurus, but nobody does anything about it. It's too bad because I. colberti could really stand a redescription, if only because the measurements and the figures in the scale bars of Jain and Bandyopadhyay (1997) don't quite jive. Hey, it's only one of the more complete titanosaurs, and it also happens to have had very unusual proportions, only we can't be sure exactly *how* unusual they were. |
| Perhaps *this* unusual; perhaps more, perhaps less. Illustration by Dmitri Bogdanov, found on Wikimedia Commons. |
{kind=link}
The partial skeleton that became the type of Isisaurus colberti was excavated 1984–1986 by the Geological Studies Unit of the Indian Statistical Institute (ISI; Kolkata) from the Lameta Formation at Dongargaon, Maharashtra, only about 16 km (10 mi) southeast of one of the earlier Lameta localities of Pisdura. The site is dominated by variegated clays with some thin partings of coarser and/or more consolidated sedimentary rocks, and is thought to represent fresh water deposition. Snails, bivalves, ostracodes, fish, turtles, crocodilians, and dinosaur eggshells are also reported from the site, although not all from the same level. The I. colberti skeleton was found in green clay, smeared out such that each bone was within a meter or so of its nearest neighbor, but the whole thing stretched over a number of meters. Together, there were nine cervicals, seven dorsals, fourteen ribs, the sacrum, sixteen caudals, nine chevrons, the left scapula and coracoid, most of the pelvis except for the left ischium, and the left humerus and ulna, catalogued as ISI R 335/1 through 65 (Jain and Bandyopadhyay 1997). Jain and Bandyopadhyay (1997) assigned it to Titanosaurus as new species T. colberti, for Edwin "Ned" Colbert. Wilson and Upchurch (2003) then moved it to its own genus, Isisaurus, honoring the Indian Statistical Institute, giving us something like "Ned Colbert's Indian Statistical Institute lizard". Incidentally, although the eye may read "Isis-saurus", you should instead think "Isi-saurus" (or, I suppose, "eye-ess-eye-saurus" if you're feeling pedantic).
As I mentioned, I. colberti was doing something very strange with its proportions. The cervicals are very short in length while also having tall neural arches, making for an unusually short and deep neck with a lot of superstructure. The cervical ribs were short and stout, unlike the cervical ribs of some sauropods that run beneath several cervicals. Exaggeration of the neural arches continues into the dorsals, which have very broad and tall neural arches. Meanwhile, the humerus is 148 cm long (45 in), quite a bit longer than the 108-cm-long (33 in) scapula and the 80-cm-long (24 in) ulna, which is otherwise notable for its distinctive triangular cross-section. The overall impression is that the neck, shoulders, and arm segments were drawn up and installed by different crews which had no contact with each other, or perhaps it had been planning on a career as a staircase or very short bridge. You've got this short thick neck, undersized shoulder girdle (which has a kind of chunky saltasaur-like appearance to it), elongate upper arm, and stocky forearm. One wonders how the hind limbs looked. The ilia are restored as flat elements pitched up slightly anteriorly, and the pubis is elongate and close to vertical. Oddly, the left ilium of the type individual is fused and coosified to the sacrum, but the right ilium isn't (Jain and Bandyopadhyay 1997).
In addition to the type skeleton, a braincase (ISI R199) was found near the I. colberti type locality in the late 1960s with some "Titanosaurus indicus"-like dorsals and caudals. Berman and Jain (1982) made the first description of this specimen. Although it cannot be definitely assigned to I. colberti, it is distinct from the Jainosaurus braincase. Similar braincases are known from Bara Simla (or Rahioli per Chatterjee and Rudra 1996) (ISI R467) and Vitakri, Pakistan (GSP-UM 7000, which Malkani 2019a considers a specimen of Saraikimasoom). All three have the occipital condyle deflected down about 120 degrees from the plane of the skull roof (Wilson et al. 2005). ISI R467 is also reportedly associated with as-yet undescribed postcrania (Chatterjee and Rudra 1996). Aside from GSP-UM 7000, potential reports of I. colberti in Pakistan include an ulna (Wilson et al. 2005) and the material Malkani formerly described as Sulaimanisaurus gingerichi, which he now considers a synonym of I. colberti (Malkani 2019a).
Phylogenetically speaking, I. colberti tends to show up in the vicinity of Rapetosaurus and sometimes Tapuiasaurus (González Riga et al. 2016, 2018; Carballido et al. 2017; Carvalho et al. 2017; Fernández-Baldor et al. 2017; Mannion et al. 2017; Tykoski and Fiorillo 2017; Averianov and Efimov 2018; Sallam et al. 2018; Simón et al. 2018; Filippi et al. 2019), except when it doesn't (Gallina and Otero 2015; Bandeira et al. 2016; França et al. 2016; Gorscak et al. 2017; some analyses in Gorscak and O'Connor 2019), in which case all bets are off as to the company it keeps. Of course, where exactly the Isisaurus/Rapetosaurus duo is found in any given tree varies widely.
Jainosaurus septentrionalis
The nexus of the issues with the Lameta sauropods is the "Sauropod Bed" on the west slope of Bara Simla, which is the source of the type material of Titanosaurus indicus, the type braincase of Jainosaurus septentrionalis, and the postcranial material that at various times has been thought to belong to one or the other, or both (or both in one when they've been thought to be synonyms). We'll leave the detailed history of T. indicus for another time, which means we skip ahead to October 1917, the beginning of the excavation of the Jainosaurus-Titanosaurus complex. These excavations came up with material including a braincase, three caudals, twenty-eight fragments of rib shafts, chevrons, two scapulae, a sternal plate, a humerus, a radius, an ulna, a tibia, and a fibula (Huene and Matley 1933; Wilson and Upchurch 2003).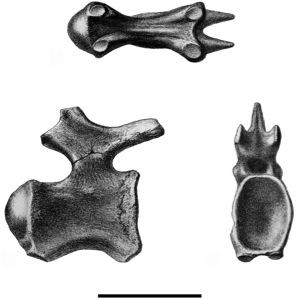 |
| A type caudal of Titanosaurus indicus, which is to blame for all of this. The type caudals appear to be lost, perhaps because they know they're in trouble. Modified slightly from Falconer (1868), via Jeff Wilson's "Introduction to Titanosauria". |
Matley (1921) was of the opinion that the sauropod bones belonged to one individual. Friedrich von Huene, who studied the material when it was in the British Museum of Natural History, came to different conclusions, such that by Huene and Matley (1933) there were at least three individuals from at least two genera. Huene and Matley (1933) placed two of the caudals, a chevron, and the hindlimb bones in Titanosaurus indicus, and assigned the rest of the material to a new species of Antarctosaurus, A. septentrionalis, with at least two individuals represented. Nearly 90 years later it's hard to understand why they should have chosen to place the new species in a genus otherwise only known from South America, as opposed to Titanosaurus or a new genus (or Argyrosaurus or Laplatasaurus for that matter), but it may have had something to do with A. wichmannianus having the only other known titanosaur braincase at the time (Wilson and Upchurch 2003). The action was consistent, though, fitting right in with having numerous South American species of Titanosaurus, or sinking Titanosaurus madagascariensis into the South American genus Laplatasaurus and then assigning some Indian fossils to Laplatasaurus madagascariensis.
There are several basic strategies that can be employed for the Bara Simla Sauropod Bed titanosaur material:
- The strict approach: J. septentrionalis and T. indicus are limited to their type specimens, with everything else to be determined on a case-by-case basis (ideally once specimens with better association are found);
- The Jainosaurus approach: the Bara Simla material excavated from October 1917 on is basically one individual animal which should be considered J. septentrionalis, with the original T. indicus holotype set aside as being impossible to assess one way or the other (and dubious anyway) (Wilson and Upchurch 2003; Wilson et al. 2009, 2011);
- The Titanosaurus approach: the Bara Simla material is basically one animal which should be considered T. indicus (Chatterjee and Rudra 1996; Jain and Bandyopadhyay 1997);
- The von Huene approach: some of the Bara Simla material is J. septentrionalis and some is T. indicus, divided by characteristics of size and robustness. Malkani's approach is similar in spirit to the von Huene approach, except with different taxa: T. indicus and the type braincase of J. septentrionalis are probably the same as Pakisaurus balochistani, but some of the Bara Simla material belongs to smaller, stockier gspsaurs.
In essence, the sauropods of Huene and Matley (1933) suffer from a lack of information. Von Huene was operating at a time without a viable theory of plate tectonics, and without having any idea that the Cretaceous unfolded over nearly 80 million years. (In 1915, American Museum of Natural History curator William Diller Matthew published a diagram that placed the end of the Mesozoic three million years ago and gave the entire age of the Mesozoic as nine million years [Norell et al. 1995].) He had no reason to suspect that by 2019 there would be more than a hundred "titanosaur" species, almost all of which were equipped with procoelous caudals just like T. indicus, but which display a dizzying variety of skeletal proportions. As far as he knew there were only a few genera and species of titanosaurs, existing over a few million years at most. This isn't to completely absolve him of his questionable taxonomic decisions, but it does place him in his time.
My opinion? I think it's reasonable that the arm, shoulder, and rib material go together (c.f. Wilson and Upchurch 2003, Wilson et al. 2009). It's also certainly possible that the Bara Simla Sauropod Bed includes multiple sauropod species, as suggested by von Huene and Malkani, but I put nothing past titanosaurs when it comes to their anatomy. It's hard to say how T. indicus fits in because there's no way of knowing how the type material relates to the other bones from the Sauropod Bed, and because we could really use a few good Lameta titanosaur tails.
Anyway, back to J. septentrionalis proper. The original name is a bit of a joke; as mentioned, this was in the days before plate tectonics, so it was assumed that Cretaceous India was about where it is today. "Antarctosaurus" means "southern lizard", but the new species was from the Northern Hemisphere, so Huene and Matley gave it a species name meaning "of the north". Therefore, Antarctosaurus septentrionalis basically means "northern southern lizard" (Wilson et al. 2009). Of course, the wordplay of the species name makes no sense with the species moved out of Antarctosaurus, but them's the breaks. Ironically enough, we now know that it was in the Southern Hemisphere at the time it existed. At any rate, Hunt et al. (1995) moved "A." septentrionalis to a new genus, Jainosaurus, named for Indian paleontologist Sohan Jain, so now it's just "Sohan Jain's northern lizard". The type specimen is the braincase GSI K27/497 (Wilson et al. 2009). Aside from not having the 120 degree angle for the occipital condyle seen in the possible Isisaurus braincases, the braincase is also noted for its well-developed parietal ridge (Berman and Jain 1982; Wilson et al. 2005). Should the postcranial material also belong, this species would combine a very long and slender scapula with a robust humerus and ulna (Wilson et al. 2009).
 |
| Several sauropod endocasts, including that of J. septentrionalis, which surely never thought it would be this much trouble after it died. Figure 6 in Knoll et al. (2015). CC-BY-4.0. |
{kind=link}
Based on comparisons with the Bara Simla postcranial material, Wilson et al. (2011) described a partial skeleton from Chhota Simla as J. cf. septentrionalis. This specimen includes a partial dorsal rib, a caudal centrum, the left humerus and partial right humerus, the right radius, and the left femur (128.8 cm or 39.3 in long), tibia, and fibula. Other bones were reportedly recovered but have gone missing, particularly vertebrae. The limb bones are useful for showing that the Bara Simla arm bones could have come from one individual. (Malkani 2019b regards the Chhota Simla material as another Pakisaurus/gspsaur mix.)
Perhaps not surprisingly for such a convoluted species, which may include anything from a single braincase to a couple of skeletons, J. septentrionalis has never been a big player in phylogenetic analyses. It appears to be close to Vahiny depereti ("Malagasy Taxon B" braincase) from Madagascar (Wilson et al. 2009).
Jiangshanosaurus lixianensis
First things first: Jiangshanosaurus is not the same as Jingshanosaurus, which is a Lower Jurassic "prosauropod". If you're worn out from Bara Simla and take nothing else from this section, take that. If you are ever in the position to name something, please make sure that there are no other organisms in the same general group with a similar name. There is no need for another Sino- or Fukui- dinosaur. Certainly don't name one thing and then give something else almost the same name. To hop back to India for a second, between the two of them, von Huene and Matley should have been able to come up with something other than Indosaurus and Indosuchus.J. lixianensis was described in 2001 by Feng et al. The genus name refers to Jiangshan County in Zhejiang Province, and the species name refers to Lixian Village, where the type and only known specimen was found, giving us something along the lines of "Jiangshan County lizard from Lixian". The type specimen, M1322 in the Zhejiang Natural Museum, consists of most of a left scapulocoracoid, five dorsal vertebrae, three caudals, and parts of both pubis bones, both ischia, and a femur. The type horizon is reported to be the Albian-age lower Jinhua Formation (Feng et al. 2001).
At least one of the caudals is slightly procoelous. This, along with the neural arches being on the anterior part of the caudal centra, and the absence of pleurocoels on the caudals, are used to assign the species to the titanosaurs. Both ends of the femur are missing, but a 84-cm-long (26 in) segment of the shaft is present. Feng et al. (2001) described the scapulocoracoid as similar in size and shape to that of North Horn Formation Alamosaurus sanjuanensis. It might be the perspective of the illustrations, but the resemblance seems a bit overstated to me.
Be that as it may, J. lixianensis has made it into a surprising number of phylogenetic analyses for something amassing about a dozen bones, and several of them find it adjacent to Alamosaurus and positioned as a very derived titanosaur (Mannion et al. 2013, 2017; Poropat et al. 2016; Averianov and Efimov 2018; Gonzàlez Riga et al. 2018). The latter point is surprising given J. lixianensis is one of the few Early-Cretaceous-aged forms to consistently turn up in Titanosauria. However, a few other recent references regard it as much more basal within Titanosauria (Averianov and Sues 2017; Sallam et al. 2018; Gorscak and O'Connor 2019). [Added 2019/09/01: Even more recently, Mannion et al. (2019) published three analyses with different results: a euhelopodid outside of Titanosauria; cavorting within a poorly resolved cloud of euhelopodids, basal titanosaurians, and unaffiliated somphospondyls; or a lithostrotian. Their restudy of the anatomy indicates that the lithostrotian result is the least likely.]
There's a skeletal mount, but generally I don't wish to encourage skeletal mounts for eight vertebrae, parts of the girdles, and a femoral shaft.
References
Averianov, A., and V. Efimov. 2018. The oldest titanosaurian sauropod of the Northern Hemisphere. Biological Communications 63(6):145–162. doi:10.21638/spbu03.2018.301.Averianov, A., and H.-D. Sues. 2017. Review of Cretaceous sauropod dinosaurs from central Asia. Cretaceous Research 69:184–197. doi:10.1016/j.cretres.2016.09.006.
Bandeira, K. L. N., F. Medeiros Simbras, E. Batista Machado, D. de Almeida Campos, G. R. Oliveira, and A. W. A. Kellner. 2016. A new giant Titanosauria (Dinosauria: Sauropoda) from the Late Cretaceous Bauru Group, Brazil. PLoS ONE 11(10):e0163373. doi:10.1371/journal.pone.0163373.
Berman, D. S., and S. L. Jain. 1982. The braincase of a small sauropod dinosaur (Reptilia: Saurischia) from the Upper Cretaceous Lameta Group, Central India, with review of Lameta Group localities. Annals of the Carnegie Museum 51:405–422.
Carballido, J. L., D. Pol, A. Otero, I. A. Cerda, L. Salgado, A. C. Garrido, J. Ramezani, N. R. Cúneo, and J. M. Krause. 2017. A new giant titanosaur sheds light on body mass evolution among sauropod dinosaurs. Proceedings of the Royal Society B: Biological Sciences 284(1860):20171219. doi:10.1098/rspb.2017.1219.
Carvalho, I. S., L. Salgado, R. M. Lindoso, H. I. de Araújo-Júnior, F. C. Costa Nogueir, and J. A. Soares. 2017. A new basal titanosaur (Dinosauria, Sauropoda) from the Lower Cretaceous of Brazil. Journal of South American Earth Sciences 75:74–84. doi:10.1016/j.jsames.2017.01.010.
Chatterjee, S., and D. K. Rudra. 1996. KT events in India: impact, rifting, volcanism and dinosaur extinction. Memoirs of the Queensland Museum 39:489–532.
D'Emic, M. D. 2012. The early evolution of titanosauriform sauropod dinosaurs. Zoological Journal of the Linnean Society 166(3):624–671.
Falconer, H. 1868. Notes on fossil remains found in the Valley of the Indus below Attock, and at Jubbulpoor. Pages 414–419 in C. Murchison, editor. Palaeontological memoirs and notes of the late Hugh Falconer, vol. I. Fauna Antiqua Sivalensis. Robert Hardwicke, London.
Feng T., X.-M. Kang, X.-S. Jin, F. Wei, and W.-T. Wu. 2001. A new sauropod dinosaur of Cretaceous from Jiangshan, Zhejiang Province. Vertebrata PalAsiatica 39(4):272–281.
Fernández-Baldor, F. T., J. I. Canudo, P. Huerta, M. Moreno-Azanza, and D. Montero. 2017. Europatitan eastwoodi, a new sauropod from the lower Cretaceous of Iberia in the initial radiation of somphospondylans in Laurasia. PeerJ 5:e3409. doi:10.7717/peerj.3409.
Filippi, L. S., L. Salgado, and A. C. Garrido. 2019. A new giant basal titanosaur sauropod in the Upper Cretaceous (Coniacian) of the Neuquén Basin, Argentina. Cretaceous Research 100:61–81. doi:10.1016/j.cretres.2019.03.008.
França, M. A. G., J. C. de A. Marsola, D. Riff, A. S. Hsiou, and M. C. Langer. 2016. New lower jaw and teeth referred to Maxakalisaurus topai (Titanosauria: Aeolosaurini) and their implications for the phylogeny of titanosaurid sauropods. PeerJ 4:e2054. doi:10.7717/peerj.2054.
Gallina, P. A., and A. Otero. 2015. Reassessment of Laplatasaurus araukanicus (Sauropoda: Titanosauria) from the Upper Cretaceous of Patagonia, Argentina. Ameghiniana 52:487–501. doi:10.5710/AMGH.08.06.2015.2911.
González Riga, B. J., M. C. Lamanna, L. D. Ortiz David, J. O. Calvo, and J. P. Coria. 2016. A gigantic new dinosaur from Argentina and the evolution of the sauropod hind foot. Scientific Reports 6:19165. doi:10.1038/srep19165.
Gonzàlez Riga, B. J., P. D. Mannion, S. F. Poropat, L. D. Ortiz David, and J. P. Coria. 2018. Osteology of the Late Cretaceous Argentinean sauropod dinosaur Mendozasaurus neguyelap: implications for basal titanosaur relationships. Zoological Journal of the Linnean Society 184(1):136–181. doi:10.1093/zoolinnean/zlx103.
Gorscak, E., and P. M. O’Connor. 2019. A new African titanosaurian sauropod dinosaur from the middle Cretaceous Galula Formation (Mtuka Member), Rukwa Rift Basin, southwestern Tanzania. PLoS ONE 14(2):e0211412. doi:10.1371/journal.pone.0211412.
Huene, F. von, and C. A. Matley. 1933. The Cretaceous Saurischia and Ornithischia of the central provinces of India. Palaeontologia Indica 21:1–74.
Hunt, A. P., M. G. Lockley, S. G. Lucas, and C. A. Meyer. 1995. The global sauropod fossil record. Gaia 10:261–279.
Jain, S. L., and S. Bandyopadhyay. 1997. New titanosaurid (Dinosauria: Sauropoda) from the Late Cretaceous of central India. Journal of Vertebrate Paleontology 17(1):114–136.
Knoll F., L. M. Witmer, R. C. Ridgely, F. Ortega, and J. L. Sanz. 2015. A new titanosaurian braincase from the Cretaceous "Lo Hueco" localityin Spain sheds light on neuroanatomical evolution within Titanosauria. PLoS ONE 10(10):e0138233. doi:10.1371/journal.pone.0138233.
Malkani, M. S. 2019a. Revision, discussion and diagnostic features of valid titanosaurs (Sauropoda, Dinosauria) from Indo-Pakistan landmass. Researchgate.net. Research. 132 pages.
Malkani, M. S. 2019b. Titanosaurus indicus, Titanosaurus blanfordi, Antarctosaurus septentrionalis, Jainosaurus septentrionalis, Laplatasaurus madagascariensis and Titanosaurus rahioliensis titanosaurs from India; their guide fossils affinity to recent valid titanosaurs Isisaurus, Pakisaurus, Saraikimasoom and Gspsaurus from Indo-Pakistan. Researchgate.net. Research. 40 pages.
Mannion, P. D., P. Upchurch, R. N. Barnes, and O. Mateus. 2013. Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriforms. Zoological Journal of the Linnean Society 168:98–206. doi:10.1111/zoj.12029.
Mannion, P. D., R. Allain, and O. Moine. 2017. The earliest known titanosauriform sauropod dinosaur and the evolution of Brachiosauridae. PeerJ 5:e3217. doi:10.7717/peerj.3217.
Mannion, P. D., P. Upchurch, X. Jin, and W. Zheng. 2019. New information on the Cretaceous sauropods of Zhejiang Province, China: impact on Laurasian titanosauriform phylogeny and biogeography. Royal Society Open Science 6(8):191057. doi:10.1098/rsos.191057.
Matley, C. A. 1921. On the stratigraphy, fossils and geological relationships of the Lameta Beds of Jubbulpore. Records of the Indian Geological Survey 53:142–164.
Norell, M. A., E. S. Gaffney, and L. Dingus. 1995. Discovering dinosaurs in the American Museum of Natural History. Alfred A. Knopf, Inc., New York, New York.
Poropat, S. F., P. D. Mannion, P. Upchurch, S. A. Hocknull, B. P. Kear, M. Kundrát, T. R. Tischler, T. Sloan, G. H. K. Sinapius, J. A. Elliott, and D. A. Elliott. 2016. New Australian sauropods shed light on Cretaceous dinosaur palaeobiogeography. Scientific Reports 6:article number 34467. doi:10.1038/srep34467.
Sallam, H. M., E. Gorscak, P. M. O’Connor, I. A. El-Dawoudi, S. El-Sayed, S. Saber, M. A. Kora, J. J. W. Sertich, E. R. Seiffert, and M. C. Lamanna. 2018. New Egyptian sauropod reveals Late Cretaceous dinosaur dispersal between Europe and Africa. Nature Ecology & Evolution 2:445–451. doi:10.1038/s41559-017-0455-5.
Sharma, N., R. K. Kar, A. Agarwal, and R. Kar. 2005. Fungi in dinosaurian (Isisaurus) coprolites from the Lameta Formation (Maastrichtian) and its reflection on food habit and environment. Micropaleontology 51:73–82.
Simón, E., L. Salgado, and J. O. Calvo. 2018. A new titanosaur sauropod from the Upper Cretaceous of Patagonia, Neuquén Province, Argentina. Ameghiniana 55(1):1–30. doi:10.5710/AMGH.01.08.2017.3051.
Tykoski, R. S., and A. R. Fiorillo. 2017. An articulated cervical series of Alamosaurus sanjuanensis Gilmore, 1922 (Dinosauria, Sauropoda) from Texas: new perspective on the relationships of North America's last giant sauropod. Journal of Systematic Palaeontology 15(5):339–364.
Wilson, J. A., and D. M. Mohabey. 2006. A titanosauriform (Dinosauria: Sauropoda) axis from the Lameta Formation (Upper Cretaceous: Maastrichtian) of Nand, central India, Journal of Vertebrate Paleontology 26(2):471–479.
Wilson, J. A., and P. Upchurch. 2003. A revision of Titanosaurus Lydekker (Dinosauria – Sauropoda), the first dinosaur genus with a "Gondwanan" distribution. Journal of Systematic Palaeontology 1(3):125–160.
Wilson, J. A., M. S. Malkani, and P. D. Gingerich. 2005. A sauropod braincase from the Pab Formation (Upper Cretaceous, Maastrichtian) of Balochistan, Pakistan. Gondwana Geological Magazine special volume 8:101–109.
Wilson, J., M. D'Emic, C. A. Curry Rogers, D. M. Mohabey, and S. Sen. 2009. Reassessment of the sauropod dinosaur Jainosaurus (="Antarctosaurus") septentrionalis from the Upper Cretaceous of India. Contributions from the Museum of Paleontology, University of Michigan 32(2):17–40.
Wilson, J. A., P. M. Barrett, and M. T. Carrano. 2011. An associated partial skeleton of Jainosaurus cf. septentrionalis (Dinosauria: Sauropoda) from the Late Cretaceous of Chhota Simla, central India. Palaeontology 54(5):981–998.
Thanks for a very lucid summary of the Titanosaurus indicus-Jainosaurus septentrionalis mess - Huene and Matley (1933) turned it into a dog's breakfast. I look forward to your 'take' on Titanosaurus indicus.
ReplyDeleteDo you think is it possible that all the Bara Simla "Sauropod Bed" material - including the Titanosaurus indicus and Antarctosaurus septentrionalis holotypes - belongs to a single individual?
Thank you! I think there's a chance that all the sauropod material came from one individual, but I wouldn't stake anything important on it. Without knowing where the T. indicus type caudals were found relative to Matley's specimens, about the best anyone could ever do is to put them in the same species, and that would require finding a nice associated specimen including the appropriate braincase, forelimbs, shoulder girdle, hindlimbs, and caudals.
DeleteThanks Justin. In size and morphology, the Titanosaurus indicus type caudals do compare quite well with those referred to Antarctosaurus/Jainosaurus septentrionalis.
DeleteI had thought the original (type) caudals of T. indicus (first described by Falconer) came from the "Sauropod bed" of Bara Simla. This is based on Wilson & Upchurch (2003):
"The bulk of the sauropod material was found approximately 1.2 m above the ‘Main Lameta limestone’, in the red and green marly clay of the ‘Sauropod bed’ that yielded the original remains of T. indicus... Although Matley initially believed this material belonged to a single individual, morphological features present in the sample required the presence of two taxa (T. indicus, Antarctosaurus septentrionalis) and at least three individuals (Huene & Matley 1933:4, 6–35)."
I don't doubt that the T. indicus caudals also came from the Sauropod Beds, it's just not known where they were in relation to the other bones (and for those we only have vague information as it is). There are plenty of multitaxic sauropod bonebeds elsewhere, so it's certainly not impossible that Bara Simla is too. On the other hand, it's certainly not impossible that Bara Simla is just one sauropod.
DeleteAlthough field data is lacking, the dimensions of the 'Sauropod bed' / Bara Simla titanosaur material is consistent with a single individual, as noted by Wilson & Upchurch.
ReplyDeleteMatley (1921) also thought that a single sauropod individual was preserved at the 'Sauropod bed'.
Curiously, Huene and Matley (1933) actually state "...from their mode of occurrence, the first impression was that the majority of them belonged to a single individual... but further study has shown that at least three individuals belonging to more than one genus, are represented." (p.4). As you know, this "further study" was Huene’s bizarre and idiosyncratic approach to titanosaur taxonomy, with his arbitrary sorting of associated material into individual species.
A side note but the northern southern 'joke' was actually done at least once before, namely in another, still mostly extant, dinosaur. The kaka, a New Zealand parrot, has a subspecies Nestor meridionalis meridionalis and a Nestor meridionalis septentrionalis. I guess some jokes take too long to get old among taxonomists...
ReplyDeleteBy the way, my compliments for your perseverance in going through the taxonomic mess that is Titanosauria. It takes stamina, I'm sure.
Thanks! It certainly keeps me busy!
DeleteThis page should be updated to reflect that Jiangshanosaurus is recovered outside Titanosauria as a euhelopodid by Mannion et al. (2019) (although two of the three cladograms place Jiangshanosaurus, and Euhelopodidae, in Titanosauria). Basically, Mannion et al. agree with Averianov and Sues (2017) as well as Gorscak and O'Connor (2019) that Jiangshanosaurus falls outside Lithostrotia considering its older age.
ReplyDeleteThank you for the suggestion. I'm not sure I want to go down the road of making revisions of that scale to blog posts.
DeleteI just went through the absolute headache of trying to organise the Malkani 2019 revisions into something vaguely resembling a coherent faunal list for the Lameta and Pab/Vitakri Formations and came up with something like this:
ReplyDeleteTitanosaurus indicus 1877 (including Pakisaurus balochistani 2004), Lameta and Pab/Vitakri Formations
Titanosaurus/Balochisaurus blandfordi 1879/2004/2019 (including Balochisaurus malkani 2004, "Nicksaurus razashahi" and "Sarakimasoom vitakri" [RIP]), Lameta and Pab/Vitakri Formations
Jainosaurus septentrionalis 1933/1995, Lameta Formation (I went against Malkani in keeping this species separate since there's absolutely no way to decipher exactly how many species Von Huene was working with unless we find an associated skeleton)
Isisaurus colberti 1997/2003 (including Sulaimanisaurus gingerichi 2004), Lameta and Pab/Vitakri Formations
Marisaurus jeffi 2003 (including "Gspsaurus pakistani" and "Maojandino alami"), Pab/Vitakri Formation
I can also only assume that since he didn't touch Khetranisaurus barkhani that he intended to leave it as its own species.
That leaves the Lameta Formation with Titanosaurus indicus, Titanosaurus/Balochisaurus blandfordi (depending on if you keep it in Titanosaurus), Jainosaurus septentrionalis and Isisaurus colberti, and the Pab/Vitakri with T. indicus, T./B. blandfordi, I. colberti, Marisaurus jeffi and Khetranisaurus barkhani.
Thank you!
DeleteHi Justin,
DeleteThe papers by Malkani in the references section are no longer online at ResearchGate and Malkani (2021) formally describes Balochisaurus, Nicksaurus, Sarakimasoom, Gspsaurus, Maojandino, Khetranisaurus, and Sulaimanisaurus as new genera and species. Thus, Malkani's (2006) attribution of his sauropod genera to Malkani (2004) without mentioning that the 2004 abstract isn't a valid publication under ICZN rules means Malkani (2021) should be treated as the correct authorship for Vitakridrinda, Balochisaurus, Nicksaurus, Sarakimasoom, Gspsaurus, Maojandino, Khetranisaurus, and Sulaimanisaurus.
Malkani, M. S. (2004). Saurischian dinosaurs from Late Cretaceous of Pakistan. pp. 71-73 In Hussain, S. S., and Akbar, H. D., eds., Fifth Pakistan Geological Congress, 14-15 April, Islamabad, National Geological Society of Pakistan, Pakistan Museum of Natural History (Pakistan Science Foundation), Islamabad.
Malkani, M. (2021) Jurassic-Cretaceous and Cretaceous-Paleogene Transitions and Mesozoic Vertebrates from Pakistan. Open Journal of Geology 11: 275-318. doi: 10.4236/ojg.2021.118016.